
Komen en gaan van een kleine pionier: groenknolorchis

Figuur 1. groenknolorchis (Liparis loeselii)
De groenknolorchis, een weinig opvallende maar bijzondere orchidee, komt vooral op de Waddeneilanden nog veel voor. Het is een merkwaardige pionierplant die het goed doet in vrij kalkrijke duinvalleien die zich steeds verjongen.
De groenknolorchis (Liparis loeselii), vroeger ook wel Sturmia genoemd, komt ook buiten de Wadden op verschillende plaatsen in Nederland nog in redelijke aantallen voor. In de ons omringende landen is hij echter zeer zeldzaam. Nederland draagt daarmee een grote verantwoordelijkheid voor het voortbestaan van deze soort in Europa. De groenknolorchis is opgenomen in de Europese Habitatrichtlijn en heeft in ons land de hoogste beschermingsstatus. Dit artikel beschrijft het reilen zeilen van deze soort in relatie tot de dynamiek en het beheer van zijn leefgebieden.
Habitat
De groenknolorchis komt voor in jonge duinvalleien en trilvenen. Het zijn vooral de vroege stadia in de vegetatieontwikkeling van duinvalleien en verlandende moerassen, waar vochtige, voedselarme en kalkrijke of basenrijke omstandigheden heersen. Andere bijzondere soorten van deze vegetaties zijn parnassia, knopbies, moeraskartelblad, moeraswespenorchis, vleeskleurige orchis, zomerbitterling en slanke gentiaan. Wanneer door natuurlijke processen uiteindelijk verzuring en verstruiking optreedt en de voedselrijkdom toeneemt, is de groenknolorchis één van de eerste soorten die verdwijnt.

Figuur 2. Enkele bloeiende exemplaren van de groenknolorchis in een loopduin (secundaire uitblazingsvallei). Noordvaarder, Terschelling.
In Nederland komt de groenknolorchis nog in flinke aantallen voor op de Waddeneilanden (tot meer dan 10000 ex zoals op Texel, 2013) en in Zeeland. In de vastelandsduinen is hij hier en daar te vinden waar door afplaggen een pioniermilieu is gecreëerd (zoals in het Zwanenwater) of in een door de mens aangelegde primaire duinvallei zoals het Kennemerstrand bij IJmuiden. De soort komt in laagveengebieden nog voor in de Nieuwkoopse en Ankeveense Plassen, De Wieden en de Weerribben, steeds op plaatsen waar relatief schoon en bovendien basenrijk (grond)water aanwezig is.
Voortplanting
In tegenstelling tot vele begeleidende soorten uit het zelfde milieu en vegetatiestadium heeft de groenknolorchis een relatief simpele voortplantingsstrategie. De kleine, geelgroene bloempjes zijn niet bijzonder aantrekkelijk voor insecten, en bloembezoek is dan ook nog nooit waargenomen. De stuifmeelklompjes vallen vanzelf op de stempel, zodat zelfbestuiving spontaan optreedt (Figuur 3). Dat is tevens te zien aan de vruchtzetting: bijna alle bloemen vormen een vrucht, waarin de groenknolorchis afwijkt van bijvoorbeeld de vleeskleurige orchis (Dactylorhiza incarnata) uit dezelfde habitat. De laatstgenoemde leidt insecten om de tuin door net te doen alsof hij in de bloemspoor nectar produceert, maar die spoor is leeg. Daarmee is de vleeskleurige orchis afhankelijk van insectenbezoek, zodat lang niet alle bloemen bestoven worden en een vrucht vormen.

Figuur 3. Groenknolorchis, bloeiwijze. Let op het glimmende gele bolletje bovenin de bloem; dat is het stuifmeelklompje dat reeds spontaan op de stempel gevallen is.
Doordat zelfbestuiving – en daarmee inteelt – de regel is, is de genetische variatie in een populatie van groenknolorchis relatief laag. Inteelt, die bij allerlei plantensoorten als gevolg van kleine populatiegrootte optreedt, heeft waarschijnlijk ook geen nadelige effecten meer. De nakomelingen die zg. inteeltdepressie vertonen (een afname van kieming, groei of bloei door foutjes in het DNA die bij inteelt aan het licht komen) zijn door natuurlijke selectie allang uit de populaties verwijderd. Door de beperkte genetische diversiteit reageren alle individuen in een populatie wel min of meer hetzelfde op veranderende omstandigheden: als het droog is staan ze allemaal laag in de vallei, als het nat is staan ze allemaal in een gordel hoog tegen de duinhellingen. In die zin is de soort vergelijkbaar met de éénjarige slanke gentiaan, die ook een spontane zelfbestuiver is. De geringe genetische variatie zal ook invloed hebben op de levensduur van de populatie: wanneer het milieu ongeschikt is geworden leggen nagenoeg alle individuen in een kort tijdsbestek het loodje.
Populatiedynamiek
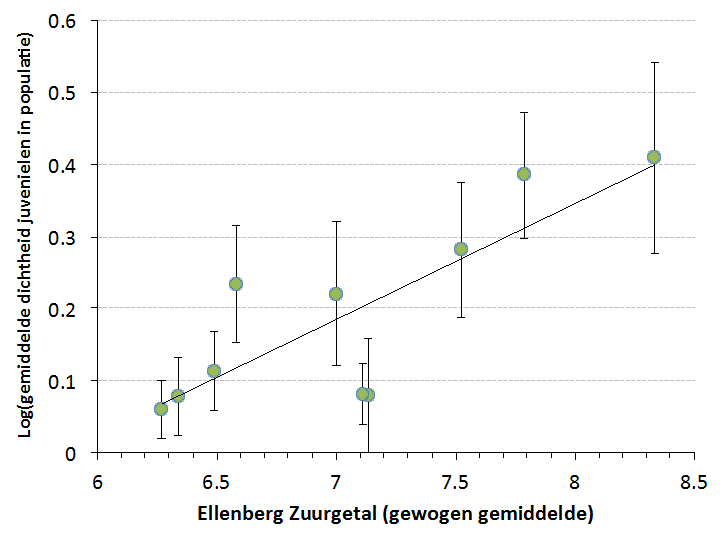
Figuur 4. Verband tussen de zuurgraad van de bodem (berekend uit de Ellenberg-indicatorgetallen voor zuurgraad van soorten in de omringende vegetatie) en de dichtheid van juvenielen van de groenknolorchis in natte duinvalleien. Het verband is statistisch significant (P=0,004): de zuurgraad verklaart 67% van de gevonden verschillen in juvenielendichtheid.
Populaties van de groenknolorchis leven gemiddeld inderdaad vrij kort. We analyseerden op grond van gegevens uit FlorBase (het bestand met inventarisatiegegevens van Floron) hoe lang de soort gemiddeld in een kilometerhok aanwezig was, en vonden dat in de meeste gevallen de levensduur van een populatie slechts tussen de 8 en 10 jaar bedroeg. Uit onderzoek van de populatie-opbouw bleek dat dit deels te maken heeft met het dichtgroeien van de vegetatie in een jonge primaire duinvallei, maar vooral met het snel zuurder worden van de bodem. Die verzuring gaat samen met de opbouw van een organische laag van afgestorven plantenresten. De dichtheid van juveniele plantjes (jonge planten met één blaadje en zonder bloemen) in een duinvalleipopulatie vertoonde een sterk positief verband met de pH (=zuurgraad) van de bodem: hoe zuurder, hoe minder verjonging (Figuur 4). Dit verband werd ook gevonden voor de totale dichtheid aan planten, maar niet voor de dichtheden van vegetatieve of reproductieve volwassen planten. Kortom: de juvenielen zijn gevoeliger voor veranderingen in het milieu dan de andere levensstadia. De gevoeligheid voor bodemverzuring betekent ook dat een beheerder weliswaar de vegetatieontwikkeling met behulp van maaien of begrazing kan afremmen, maar daarmee het verdwijnen van een pioniersoort als de groenknolorchis niet kan tegenhouden. Afplaggen werkt dan beter, omdat daardoor het kalkrijke zand (of – in laagveenterreinen – het basenrijke water) weer aan de oppervlakte komt. Door afplaggen wordt echter ook het maaiveld verlaagd, waardoor het milieu naast kalkrijker of basenrijker ook natter wordt. Uit waarnemingen van beheerders van o.m. de Inlaag Hoofdplaat (zuidoever Westerschelde), de Kreeftepolder (Texel) en de Schapenwei (Voorne’s Duin) blijkt dat de groenknolorchis daar vrij slecht tegen kan: in natte jaren werden steevast minder individuen waargenomen dan in drogere jaren.
Genetische variatie op de Waddeneilanden
De verdeling van genetische diversiteit over het landschap hangt af van de uitwisseling van genen via zaadverspreiding en verspreiding van stuifmeel. Zoals eerder gezegd is dat laatste bij groenknolorchis zeer beperkt, omdat er nagenoeg uitsluitend zelfbestuiving plaatsvindt. Daardoor vindt genenuitwisseling tussen de verschillende groeiplaatsen eigenlijk alleen door middel van zaadverspreiding plaats. Met behulp van AFLP-merkers (Amplified Fragment Length Polymorphism, een methode voor DNA-fingerprinting) hebben we dit proces voor de Waddeneilanden geprobeerd te reconstrueren. Uit het onderzoek bleek dat de populaties op grond van hun verzameling van individuele DNA-vingerafdrukken goed van elkaar te onderscheiden waren. Normaal gesproken is dit een aanwijzing voor beperkte verspreiding van zaad en stuifmeel, maar bij groenknolorchis moeten we ook rekening houden met de spontane zelfbestuiving van de soort. Theoretische modelstudies laten zien dat dit de verdeling van de genetische diversiteit sterk beïnvloedt. Toch werd er binnen de populaties nog relatief veel genetische variatie gevonden, hetgeen doet vermoeden dat ze vaak ontstaan zijn door kolonisatie van een ‘verse’ primaire duinvallei vanuit meerdere oudere populaties. Eén enkele zaadbron zou immers tot weinig variatie leiden, omdat de kolonisator nakomelingen produceert die door de zelfbestuiving allemaal zeer sterk op de moederplant lijken.
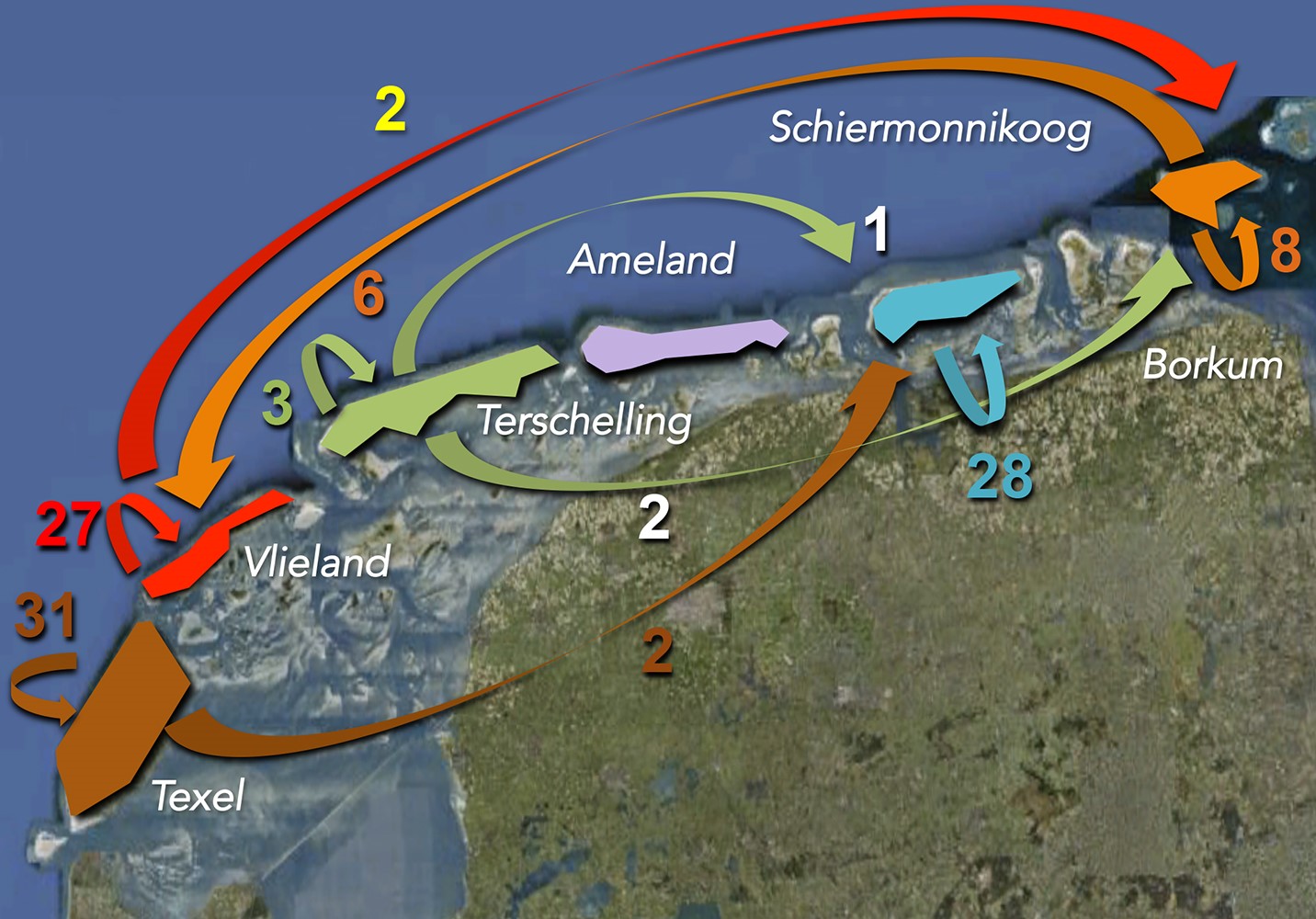
Figuur 5. Overzicht van de aantallen individuen van de groenknolorchis die op grond van hun DNA-vingerafdruk door de “toewijzingstest” aan een andere populatie dan die waar ze in voorkomen werden toegewezen. De lange pijlen betreffen verspreiding over lange afstanden, van het ene naar het andere eiland, de korte pijlen betreffen zaden die op hetzelfde eiland van de ene naar de andere populatie zijn verspreid.
De vingerafdruk-patronen in het landschap werden tevens gebruikt om de mogelijke herkomst van de individuen in recent gekoloniseerde populaties te herleiden. Dit gebeurde met zogeheten ‘toewijzingstesten’ (assignment tests). Hieruit bleek dat de meeste kolonisaties plaatsvonden vanuit nabijgelegen oudere populaties op hetzelfde eiland. Een beperkt deel van de planten was afkomstig van een ander eiland (Figuur 5). Zaadverspreiding kan dus incidenteel over zeer lange afstanden, tot wel 288 kilometer (de afstand hemelsbreed tussen Vlieland en Borkum), plaatsvinden. De vraag is of dit alleen via de wind gebeurt, of ook regelmatig via de laarzen of schoenen van duinvallei-onderzoekers… We moeten er overigens niet van uitgaan dat nieuwe duinvalleien altijd snel ‘gevonden’ zullen worden door de zaden van de groenknolorchis: in Cupido’s Polder op de Boschplaat van Terschelling bestaat al meer dan tien jaar geschikt habitat voor de soort, maar werden de eerste individuen pas enkele jaren geleden waargenomen.
Levensvatbaarheid
Om de levensvatbaarheid van de groenknolorchis op de lange termijn te garanderen is het vanwege de hoge dynamiek en korte levensduur van individuele populaties noodzakelijk dat er regelmatig nieuw habitat ontstaat. In een netwerk van populaties (een zogenaamde ‘metapopulatie’) is het niet erg dat er populaties verdwijnen, zolang er maar voldoende nieuwe bijkomen door kolonisatie van nieuwe duinvalleien (of, in het binnenland, nieuwe verlandende petgaten). In duingebieden gebeurt dit laatste door de vorming van primaire duinvalleien, wat op de Waddeneilanden nog steeds veel voorkomt, zoals op de Noordsvaarder op Terschelling en De Hors op Texel. In de vastelandsduinen vindt echter geen primaire duinvalleivorming meer plaats. Met een dynamischer kustbeheer, waar gaten in de zeereep mogen ontstaan en overstromingsvlakten (‘washovers’) gevormd mogen worden, ontstaat wellicht ook in dit gebied nieuw habitat voor de groenknolorchis en zijn begeleiders. Ook het ontstaan van strandvlaktes zoals het Kennemerstrand (na verlenging van de Zuidpier bij IJmuiden) biedt goede perspectieven, maar we moeten ons wel realiseren dat dergelijke strandvlaktes relatief kort geschikt zijn voor de zeldzame soorten van de jongste ontwikkelingsstadia. Op kalkrijke bodem of in gebieden met kalkrijke kwel kan de geschiktheid met behulp van maaibeheer echter vrij lang gerekt worden, zoals bij IJmuiden ook fraai zichtbaar is. In laagveengebieden verdwijnt habitat van de groenknolorchis door het dichtgroeien, verdrogen en verzuren van trilvenen. Dit kan alleen worden tegengegaan door regelmatig nieuwe pet- of trekgaten te graven op locaties met een kansrijke aanvoer van basenrijk (kwel)water. Lokaal is ook succes geboekt door het eerder genoemde afplaggen van veenmosrietlanden, waardoor het bufferende water weer toe kan treden tot het wortelbereik en de verzuring afneemt.
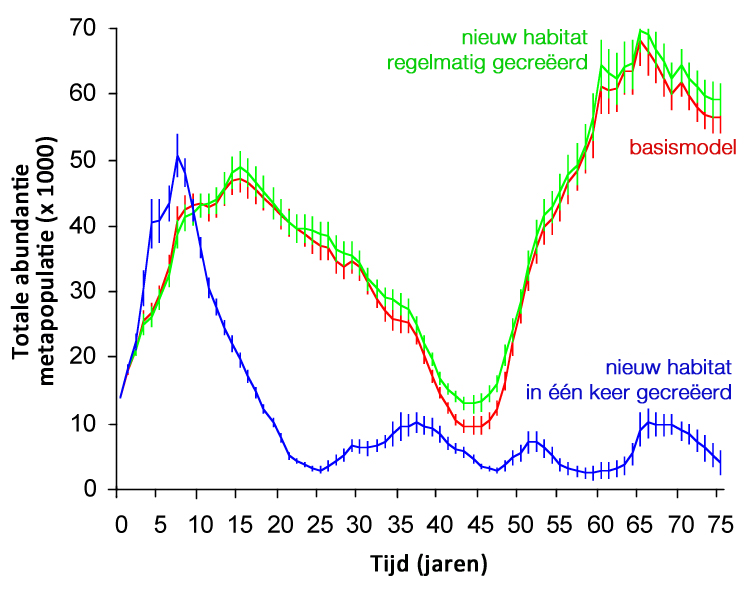
Figuur 6. Resultaten van een simulatiemodel gebaseerd op de demografie en verspreiding van de huidige populaties van de groenknolorchis, en op de verwachte ontwikkeling van habitat op grond van natuurbeleids- en beheerplannen in enkele scenario’s. De grafiek laat het verschil zien in de aantalsontwikkeling van het gehele netwerk van populaties (de metapopulatie) tussen een scenario waarbij dezelfde oppervlakte nieuw habitat in één keer wordt gecreëerd, bijv. na een grote EU-subsidie, of in de vorm van het regelmatig realiseren van kleinere oppervlakten.
Bij het creëren van nieuw habitat voor pioniersoorten moet verstandig worden omgesprongen met subsidies. Uit ons metapopulatiemodel voor de groenknolorchis bleek bijvoorbeeld dat een scenario waarbij veel habitat ineens wordt gecreëerd (bijvoorbeeld na een grote EU-subsidie voor het graven van een grote oppervlakte aan petgaten) weliswaar tot een piek in de grootte van de het hele netwerk van populaties leidt, maar dat deze metapopulatie vervolgens instort als gevolg van het feit dat er overal tegelijk een tekort aan geschikt habitat optreedt (Figuur 6). Het is dus beter om het creëren van geschikt habitat in de tijd uit te spreiden, zodat de extincties en kolonisaties in de metapopulaties van allerlei soorten meer in balans zijn.
Verantwoording
Dit verhaal kon alleen maar geschreven worden dankzij de inzet, kennis en hulp van een groot aantal mensen. Een viertal studenten van de Universiteit van Amsterdam deed onder begeleiding van Gerard Oostermeijer, Annemieke Kooijman en Patrick Meirmans hun afstudeeronderzoek aan de Groenknolorchis: Yorike Hartman (populatie-ecologie en metapopulatiemodel), Cathy Liu en Sascha van der Meer (populatiegenetica) en Annelies van der Craats (relaties bodem en vegetatie met leeftijd duinvalleien). Aan de Rijksuniversiteit Groningen deed Rohani Shahrudin haar promotieonderzoek aan (onder meer) de Groenknolorchis, onder begeleiding van Ab Grootjans, Annemieke Kooijman en Gerard Oostermeijer. Rohani verdedigt op 19 mei 2014 aan de RUG haar proefschrift, getiteld “Do we really need management to preserve pioneer stages in wet dune slacks?” Ook zijn we veel dank verschuldigd aan alle terreinbeheerders die hun kennis over de groenknolorchis en het beheer van duinvalleien en laagveengebieden met ons deelden en toestemming gaven om in hun terreinen te werken. Een deel van het onderzoek werd mogelijk gemaakt door een subsidie aan het OBN-deskundigenteam Duin- en Kustlandschap.


kan het zijn dat ik er één zag bij het zwanenwater bij Petten/
dd 13 juni 2021
ja zeker!